Ogni Giorno, ogni minuto, in ciascuna
delle nostre cellule ha luogo una scena che potrebbe essere tratta da un film
di Indiana Jones. Una innocente proteina si aggira per la cellula cercando di
fare il proprio lavoro, ma un attimo dopo viene marchiata per essere distrutta
e risucchiata in un antro oscuro, dove in men che non si dica viene ridotta
a pezzettini. A differenza di quanto succede a Indiana Jones, non c'e' scampo
per la povera proteina. Nella camera della morte essa viene stirata come in
una tortura medievale e fatta passare sotto una serie di coltelli enzimatici
che la riducono a brandelli. Pochi secondi più tardi quello che resta della
proteina emerge dal tunnel, giusto in tempo per essere afferrato e ulteriormente
maciullato da enzimi più semplici.
Si direbbe che questo dramma cellulare sia un fatto trascurabile, ma scienziati
di diversi laboratori stanno scoprendo che queste "camere della morte" molecolari,
chiamate proteasomi, in realtà hanno un ruolo fondamentale in numerosi processi
cellulari. Una tipica cellula dell'organismo possiede all'incirca 30.000 proteasomi.
Quando questi non funzionano a dovere, diverse malattie possono essere in agguato.
Alcuni virus, come quello umano dell'immunodeficienza acquisita (HIV), hanno
sviluppato sistemi in grado di manipolare il processo di degradazione delle
proteine per i loro scopi. Possiamo prevedere che molti farmaci di nuova generazione
per la cura del cancro e di altre patologie consisteranno in composti chimici
che agiscono sui proteasomi e sui meccanismi che trasportano le proteine al
loro interno. Molte industrie biofarmaceutiche hanno allo studio molecole che
inibiscono l'azione dei proteasomi; due di questi potenziali farmaci sono già
in fase di sperimentazione clinica sull'uomo.
Il gioco di costruzioni più serio del mondo
Le proteine costituiscono la vera e propria essenza delle cellule. Alcune
di esse funzionano come enzimi, gli operai molecolari che si incaricano di tulle
le reazioni chimiche necessarie alla vita. Il tipo di proteine che una cellula
produce dipende da quali dei suoi geni sono attivi in un determinato momento.
I geni contengono il codice che decide come i 20 "mattoni" fondamentali delle
proteine, gli amminoacidi, vengono assemblati in catene nelle combinazioni più
varie. Le catene a loro volta si avvolgono in eliche per formare i vari tipi
di proteine, ciascuna delle quali è caratterizzata da una specifica funzione
dettata dalla propria forma e dalle caratteristiche chimiche.
Che cosa succede quando una proteina non è più necessaria oppure assume un avvolgimento
errato? Per anni, si è ritenuto che gran parte delle proteine venisse degradata
nei lisosomi, sacche piene di enzimi digestivi che si ritrovano in quasi tutte
le cellule dell'organismo. Tuttavia, nei primi anni settanta, uno di noi (Goldberg)
scoprì che anche le cellule prive di lisosomi, come i batteri e i globuli rossi
immaturi, sono in grado di distruggere rapidamente le proteine anomale. Questo
processo inoltre richiede energia, al contrario di altri fenomeni degradativi.
Goldberg e colleghi riuscirono a riprodurre sperimentalmente le reazioni di
degradazione dipendenti dall'energia, permettendo ad altri gruppi di ricerca,
tra la fine degli anni settanta e l'inizio degli anni ottanta, di identificare
gli enzimi responsabili di questo processo. Nel 1988, infine, due gruppi, l'uno
guidato da Goldberg e l'altro da Martin C. Rechsteiner dell'Università dello
Utah, scoprirono che le proteine vengono demolite da grandi complessi multienzimatici
che il gruppo di Goldberg battezzò proteasomi.
Il nome deriva dal fatto che questi complessi contengono numerose proteasi,
enzimi che tagliano a pezzi le proteine. I proteasomi sono però 100 volte più
grandi e più complessi delle proteasi. Quando una proteina viene depositata
all'ingresso di un proteasoma, è trasportata all'interno della particella e
infine smontata negli amminoacidi che la compongono, i quali a loro volta potranno
essere riassemblati per formare altre proteine, come in un gioco di costruzioni.
La maggior parte delle proteine viene sostituita ogni pochi giorni, anche in
cellule che si dividono raramente, come quelle del fegato o del sistema nervoso.
Inoltre proteine diverse vengono degradate a velocità diverse: alcune hanno
una vita media di 20 minuti, altre possono durare per giorni o settimane. La
velocità di degradazione varia poi in modo drastico in funzione dei cambiamenti
dell'organismo.
Anche se tutto ciò può sembrare uno spreco, in realtà ha scopi ben precisi.
La degradazione di un enzima o di una proteina regolatoria, per esempio, è un
meccanismo comune con cui le cellule rallentano o interrompono una reazione
biochimica. D'altra parte la rimozione di un'importante proteina inibitoria
può aprire la strada a molti processi cellulari. L'eliminazione rapida di proteine
regolatorie è molto importante nel determinare l'esatta transizione fra i diversi
stadi del ciclo che porta alla divisione
cellulare.
La degradazione delle proteine ha un ruolo cruciale anche nella regolazione
del metabolismo generale. In caso di necessità, per esempio durante il digiuno
o in situazioni patologiche, il sistema dei proteasomi diventa più attivo nei
muscoli, fornendo amminoacidi che l'organismo può convertire in glucosio da
bruciare. E proprio l'eccesso di demolizione proteica a causare il deterioramento
muscolare e la debolezza tipici dei casi di denutrizione e negli stadi avanzati
di tumori o AIDS, o di diabete non curato.
Anche il sistema immunitario, nella sua costante lotta per eliminare delle cellule
infettate da virus o quella cancerose, utilizza i proteasomi per "etichettare"
queste cellule pericolose. Sebbene la maggior parte delle proteine cellulari
sia in genere degradata completamente fino allo stadio di singoli amminoacidi,
alcuni frammenti composti da otto o dieci amminoacidi vengono liberati dai proteasomi,
catturati e infine esposti sulla superficie cellulare, dove il sistema immunitario
può controllare se si tratta di proteine normali o anomale. In situazioni patologiche
e in alcuni tessuti come la milza e i linfonodi sono prodotti particolari tipi
di proteasomi, gli immunoproteasomi, che hanno proprio il compito di aumentare
l'efficienza di questo meccanismo di sorveglianza.
La distruzione delle proteine operata dai proteasomi è anche un importante sistema
di controllo di qualità che evita l'accumulo di proteine potenzialmente tossiche.
Sia i batteri sia le cellule dei mammiferi distruggono selettivamente le proteine
che hanno una conformazione fortemente anomala a causa di mutazioni, di errori
nella sintesi o di danni subiti.
La degradazione delle proteine anomale ha un ruolo importante in molte malattie
genetiche umane. In diverse anemie ereditarie, un gene mutante causa la produzione
di proteine di emoglobina anomale, che non si ripiegano correttamente e vengono
distrutte dai proteasomi poco dopo la sintesi. Analogamente, la fibrosi cistica
è causata da una mutazione del gene per una proteina-canale che trasporta il
cloro attraverso la membrana cellulare esterna. Dato che i canali del cloro
mutanti sono leggermente deformati, i proteasomi li degradano prima che possano
raggiungere la membrana cellulare. Il muco viscoso che si viene a creare nei
polmoni e in altri organi delle persone affette da fibrosi cistica è una conseguenza
della mancanza dei normali canali del cloro. Viceversa, l'incapacità dei proteasomi
di degradare alcune proteine potrebbe contribuire a patologie anche gravi. Si
sta scoprendo, per esempio, che ammassi di proteine mal ripiegate si accumulano
insieme con i proteasomi in alcuni neuroni nel cervello delle persone colpite
da malattie neurodegenerative. Il motivo per cui i neuroni colpiti di questi
pazienti non riescono a degradare le proteine anomale è oggetto di studi sempre
più numerosi.
Nel ventre della balena
Dal punto di vista di una proteina, i proteasomi appaiono come strutture gigantesche.
Mentre la taglia media di una proteina è compresa fra 40.000 e 80.000 dalton
(ossia fra 40.000 e 80.000 volte il peso molecolare di un atomo di idrogeno),
la maggior parte dei proteasomi che si trovano negli organismi superiori arriva
a pesare ben 2 milioni di dalton. A metà degli anni novanta un gruppo di ricercatori
guidato da Wolfgang Baumeister e Robert Huber del Max-Planck-Institut für Biochemie
a Martinsried, in Germania, grazie a tecniche di diffrazione di raggi X e di
microscopia elettronica, determinò l'architettura molecolare dei proteasomi.
Ognuno di essi e formato da una particella principale a forma di tubo e da una
o due particelle più piccole ad azione regolatoria, posizionate come tappi a
una o a entrambe le estremità. La particella principale è formata da quattro
anelli adiacenti - ognuno a sua volta composto da sette subunità - che racchiudono
un canale centrale, il quale rappresenta il tubo digerente del proteasoma. I
due anelli più esterni funzionano probabilmente da cancelli, impedendo che le
proteine possano cadere per errore nella camera di distruzione. Analogamente,
si ipotizza che anche le particelle che fungono da tappo siano guardiani estremamente
selettivi posti all'entrata della particella principale. Queste particelle regolatorie
riconoscono e legano le proteine destinate alla distruzione; utilizzano poi
energia per svolgere la struttura di queste proteine e introdurle nella particella
principale, dove sono tagliate in pezzi di varie dimensioni.
Diversi gruppi di ricerca sono riusciti a isolare o sintetizzare composti che
inibiscono selettivamente il proteasoma senza agire su altri enzimi cellulari,
evitando il rischio di effetti collaterali. Servendosi di questi inibitori gli
scienziati hanno potuto decifrare i meccanismi di funzionamento dei proteasomi.
In alte dosi, questi inibitori uccidono la cellula, cosa che non sorprende,
considerando le tante funzioni in cui i proteasomi hanno un ruolo critico. Tuttavia
è interessante il fatto che le cellule tumorali sembrino più suscettibili delle
cellule normali a questi effetti letali. Un inibitore dei proteasomi, in fase
di sperimentazione presso la Millennium Pharmaceuticals di Cambridge, nel Massachusetts,
è stato provato con successo per quanto riguarda la sicurezza nell'uomo, e nel
prossimo inverno sono previste sperimentazioni cliniche per valutarne l'efficacia
contro diversi tumori. Un altro inibitore dei proteasomi è attualmente testato
sull'uomo in vista di un suo utilizzo per la cura dell'ictus e dell'infarto
del miocardio.
Il bacio della morte
Il proteasoma non sceglie a caso le proteine da distruggere: è la cellula
a indicare quali siano destinate alla morte. Gli scienziati hanno scoperto che,
prima di venire distrutte, le proteine sono in genere "marchiate" con un'altra
proteina, chiamata ubiquitina perché si ritrova in molti organismi diversi.
Essendo composta da soli 76 amminoacidi, l'ubiquitina è una proteina relativamente
piccola che può essere fissata a proteine più grandi formando lunghe catene.
Queste code di poliubiquitina funzionano come codici postali che indirizzano
le proteine condannate ai proteasomi. Il controllo temporale della demolizione
proteica non dipende dalla distruzione da parte dei proteasomi, ma dalla velocità
del processo di ubiquitinazione - l'aggiunta delle code di ubiquitina - che
richiede energia. Il meccanismo del legame dell'ubiquitina è stato chiarito
da Avram Hershko e Aaron Ciechanover del Technion-IsraeI Institute of Technology
di Haifa, e da Irwin Rose del Fox Chase Cancer Center di Filadelfia.
Il processo di ubiquitinazione avviene in diverse tappe che coinvolgono tre
enzimi, chiamati E1, E2 ed E3. L'enzima E1 attiva l'ubiquitina e la connette
a E2. Il terzo enzima, E3, facilita poi il trasferimento dell'ubiquitina attivata
da E2 alla proteina. Il processo si ripete fino a che un lunga catena di ubiquitine
sporge dalla proteina. La catena viene poi riconosciuta da un proteasoma, che
la ingloba.
Il mistero di come una proteina venga scelta per l'ubiquitinazione si avvolge
attorno alle proteine E3. Recentemente alcuni ricercatori, fra cui due di noi
(Elledge e Harper), hanno scoperto che esistono centinaia di proteine E3 distinte
in grado di riconoscere nella sequenza amminoacidica di altre proteine le informazioni
che ne determinano l'ubiquitinazione. In risposta all'alterazione delle condizioni
fisiologiche, dovuta per esempio a infezioni o a carenze nutritive, le cellule
possono modificare le proteine aggiungendovi gruppi fosfato. La fosforilazione
può alterare l'attività di una proteina o modificarne la capacità di legarsi
alle E3. Le proteine che non riescono ad avvolgersi o risultano danneggiate
vengono anch'esse riconosciute dalle E3, che intervengono marchiando le proteine
difettose in modo che vengano prelevate dai proteasomi. Dato che molti processi
chiave della cellula dipendono dalla stabilità delle proteine, scoprire come
questa venga regolata può svelare molti segreti della biologia.
Controllando la stabilita di proteine cruciali, gli enzimi E3 regolano molti
processi cellulari, come lo sviluppo degli arti, la risposta immunitaria, la
divisione e la comunicazione fra cellule. Perfino i ritmi circadiani e la fioritura
della piante sono sotto il controllo dagli enzimi E3. Non solo: molte proteine
E3 sono state identificate come oncogeni o antioncogèni, fornendo cosi un legame
fra l'ubiquitinazione e il processo di cancerogenesi.
Un esempio è rappresentato dall'antioncogène Von Hippel Lindau (VHL), una E3
che si ritrova spesso mutata nei tumori del rene. Il compito della VHL è quello
di ritardare la crescita cellulare, limitando lo sviluppo dei vasi sanguigni
nei tessuti. Quando questa proteina è mutata i tumori neoformati sono in grado
di generare un ricco sistema di vasi sanguigni, crescendo cosi in modo rapido.
Di recente si è scoperto che una forma ereditaria del morbo di Parkinson è dovuta
alla mutazione di un gene che codifica per un enzima E3. Per effetto della mutazione,
le proteine si accumulano in specifiche cellule del cervello e le uccidono.
I virus hanno evoluto un sistema per sfruttare l'ubiquitinazione e la degradazione
delle proteine cellulari per i loro nefandi scopi. Un esempio è dato dai virus
del papilloma umano (indicati con la sigla HPV), che possono causare verruche
genitali e portare a tumori cervicali o anali. La trasformazione in cellule
cancerose maligne è di solito bloccata dalla proteina p53, un baluardo dell'organismo
che funziona da soppressore dei tumori. Per aggirare questo guardiano cellulare
i virus del papilloma usano un trucco: producono una proteina che si lega simultaneamente
alla p53 e a un enzima E3. Il legame porta all'ubiquitinazione della p53, che
è così destinata a essere fatta a pezzi dagli enzimi dei proteasomi. In questo
modo è molto più probabile che le cellule indifese diventino tumorali.
Il virus HIV utilizza uno stratagemma simile per distruggere la proteina CD4,
che si trova sulla superficie cellulare. Questa proteina è necessaria all'HIV
per infettare le cellule, ma in seguito la sua presenza interferisce con la
produzione di nuove particelle virali. Dato che si lega alla proteina gpl60,
che sporge dalla superficie del virus, CD4 funziona da "ormeggio" per l'HIV,
permettendogli di infettare i linfociti T. Tuttavia, CD4 rappresenta un problema
quando l'HIV comincia a replicarsi nelle cellule appena infettate, perché aderisce
alle proteine gp160 neosintetizzate impedendo loro di assemblarsi con le altre
proteine che compongono le particelle virali. Per aggirare questo ostacolo l'HIV
produce una proteina chiamata Vpu, grazie alla quale la CD4 viene immessa nella
corsia preferenziale verso la distruzione. Vpu si lega sia a CD4 sia a un complesso
che contiene l'enzima E3, causando l'ubiquitinazione della CD4 e il suo annientamento
nei proteasomi.
Gli studi indicano che gli enzimi E3 sono un potenziale bersaglio per i farmaci
del futuro. Dato che ciascun E3 è responsabile della distruzione di poche proteine,
i composti in grado di inibire in modo specifico i diversi E3 sarebbero molto
selettivi. Di recente l'identificazione di famiglie di enzimi E3 ha aperto nuove
strade per la messa a punto di tali farmaci.
Si tratta di sviluppi che aumentano le conoscenze dei meccanismi di regolazione
fondamentali nella biologia umana. Più cose si scoprono sui proteasomi e sui
meccanismi di degradazione proteica, più ci si rende conto del legame che esiste
fra la vita degli organismi e la morte delle proteine.
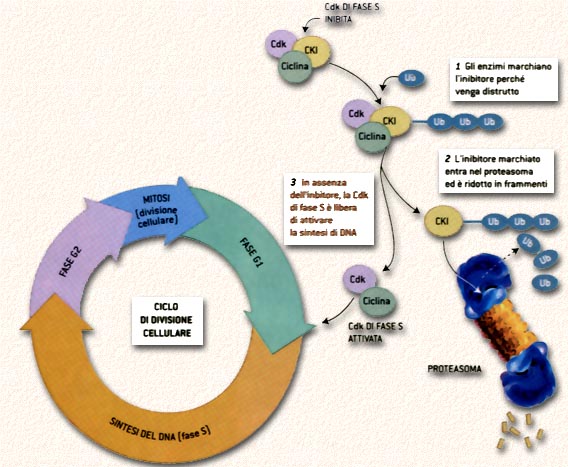
Divisione cellulare e distruzione delle proteine
Un ottimo esempio di come la capacita di una cellula
di degradare le proteine sia fondamentale per la sopravvivenza e lo sviluppo
viene dallo studio della divisione cellulare in Saccharomyces cerevisiae, il
comune lievito di birra. Prima di dividersi, qualunque cellula - che si tratti
di lieviti o di cellule umane - deve fare una copia del proprio DNA; e prima
di iniziare la sintesi di nuovo DNA deve attivare una particolare classe di
proteine, chiamate Cdk di fase S. Queste sono composte da due proteine, una
ciclina e una subunità Cdk. Le Cdk di fase S sono normalmente inattive, perché
legate a proteine inibitorie (chiamate CKI) prodotte durante la precedente divisione
cellulare. Per attivare le Cdk di fase S, le cellule devono liberarsi delle
proteine inibitorie inviandole a un proteasoma perché vengano degradate. Affinché
diventino un bersaglio per i proteasomi, le proteine vengono contrassegnate
dall'ubiquitina (Ub). Normalmente questo processo di etichettatura è strettamente
regolato, ma quando non funziona le cellule si dividono in modo incontrollato,
e possono dare origine ad alcuni tipi di tumore. Per controllare il processo
di ubiquitinazione e quindi la demolizione delle proteine che regolano la divisione
cellulare - le cellule utilizzano un complesso di molecole chiamato SCF. Il
complesso, che fa parte degli enzimi E3, agisce come una chiave universale provvista
di teste intercambiabili; grazie alla propria forma ogni testa si adatta in
modo specifico a una proteina.
Nel nematode Caenorhabditis elegans sono state identificate oltre 217 di queste
"teste", chiamate proteine F-Box. Diverse decine di esse sono state
finora isolate anche nelle cellule umane, e il loro numero è in continua crescita.
Il complesso SCF utilizza uno specifico insieme di teste adattatrici per riconoscere
le proteine da eliminare addizionando a esse un gruppo fosfato, in modo che
si leghino alle proteine F-Box dell'SCF. L'SCF funziona anche da intermediario,
facendo incontrare le sfortunate proteine con gli enzimi che imprimeranno loro
il marchio letale dell'ubiquitina.
Grazie alla varietà dei suoi complessi SCF, la cellula controlla in modo estremamente
fine quali e quante proteine può avere a disposizione in un determinato momento.
Fra le proteine regolate dai complessi SCF troviamo quelle che promuovono o
inibiscono il ciclo di divisione cellulare e quelle che attivano i geni.
Gli Autori:
ALFRED L. GOLDBERG, STEPHEN J. ELLEDGE e J. WADE HARPER hanno dedicato la
loro carriera allo studio della degradazione delle proteine e al legame di questi
processi con le malattie. Goldberg insegna biologia cellulare alla Harvard Medical
School. Nel 1998 è stato insignito del premio Novartis-Drew per la ricerca biomedica.
Elledge insegna biochimica al Baylor College of Medicine ed è ricercatore dell'Horward
Hughes Medical Institute. Nel 1994 ha ricevuto il premio Michael E. DeBakey
for Research Excellence. Harper è professore nei Dipartimenti di biochimica
e biologia molecolare dello stesso istituto. Nel 2000 ha ricevuto la Vallee
Visiting Professorship presso l'Università di Oxford.
BIBLIOGRAFIA:
KOEPP DEANNA M., HARPER J. WADE ed ELLEDGE STEPHEN J., How the Cyclin Became
a Cyclin: Regulated Proteolysis in the Cell Cycle in «Cell»,97,n.
4,14 maggio 1999.
BORMAN STU, Instricacies of the Proteasome in «Chemical and Engineering
News»,78,n. 12,20 marzo 2000.
GOLDEBRG ALFRED L., Probing the Proteasome Pathway in «Nature
Biotechnology»,18,n. 5,18 maggio 2000.